
english
Original: PDF, german
Seit 1992 werden systematisch die Auswirkungen einer sehr endlichen Laufgeschwindigkeit nervlicher Impulse (ionischen Ursprungs) in Hinsicht auf mögliche, informatisch relevante Funktionen verschiedener, neuronaler Anordnungen untersucht. Unter der Maßgabe von Leitgeschwindigkeiten biologischer Systeme im Bereich von 0.5 - 120 m/s entstehen in biologischen Systemen Impulsbreiten, deren geometrische Länge bei einer angenommemenen Impulsspitze von einer zehntel Millisekunde etwa im Bereich von 50 μm - 12 mm liegt. Eine Verknüpfung von Impulswerten aus verschiedenen Quellen ist somit mitnichten statisch durchführbar. Zur Verknüpfung ist neben dem Vorhandensein der Impulse auch deren Zusammentreffen am Ort der Interferenz erforderlich. Folglich liegt die Annahme nahe, Orte von Interferenz könnten etwas mit Adressierungsprinzipien neuronaler Information zu tun haben.
Leitgeschwindigkeit und Ortsauflösung
Obwohl in der Medizin bekannt (Klassifikationen nach Erlanger/Gasser 1939 sowie nach Lloyd/Hunt 1943 [1]), blieb die Laufgeschwindigkeit nervaler Impulse von der AI-Forschung vierzig Jahre lang vernachlässigt. Offenbar verleitete die Modellierung nervaler Eigenschaften mit verzögerungsfreien, elektrischen Leitbahnen zu diesem Mißgeschick. Idealisierte, elektrische Leitungen verschleiern, daß die Funktion eines logischen Bauelements nur gegeben ist, wenn die Signaldauer d sehr groß im Vergleich zu systemimmanenten Verzögerungszeiten T der Leitbahnen ist. Genau das aber ist bei biologischen Systemen nicht der Fall, geometrische Impulslänge und Laufzeiten der Anordnung liegen in derselben Größenordnung: d ≈ T. Wie zu zeigen sein wird, hat der entstehende Logiktyp mit dem aus der Computertechnik bekannten, statischen Typ nichts gemein.
Tabelle 1: Ausgewählte Leitgeschwindigkeiten und geometrische Breite eines 0,1ms-Impulses
| Faser-Durchmesser [μm] | Typ nach Erlanger/ Gasser | Typ nach Lloyd/ Hunt | Leitgeschwin- digkeit [m/s] |
Funktion und Vorkommen z.B. | geometrische Breite eines 0,1ms-Impulses [mm] |
| 12-20 | Aα | I | 70...120 | Versorgung quergestreifter Muskulatur | 7...12 |
| 6...12 | Aβ | II | 30...70 | Hautafferenzen für Berührung und Druck | 3...7 |
| 3...6 | Aγ | - | 15...30 | motorische Versorgung zu Muskelspindeln | 1,5...3 |
| 2...5 | Aδ | III | 12...30 | Hautafferenzen, Drucksensibilität | 1,2...3 |
| 1...3 | B | - | 3...15 | Symphatisch, präganglionär | 0,3...1,5 |
| 0,4...1,2 | C | IV | 0,5...2 | marklose Fasern, Schmerz der Haut | 50...200μm |
Die Funktion eines Gatters wird nicht mehr nur von dessen Logiktyp bestimmt. In die logische Funktion geht zusätzlich der Ort ein. Nur im Ort einer Interferenz erscheint eine Ausgabe. Die Relevanz dieser Aussage wird bei einem Blick auf die Tabelle deutlich. Es ist zu beachten, daß insbesondere die dünnsten, marklosen (d.h. nichtisolierten) Fasern sehr niedrige Leitgeschwindigkeiten verursachen. Feine Fasern bilden den sog. Filz des Nervengeflechts. Die Annahme einer Impulsdauer von 0,1ms entspricht dem steilsten Teil eines Peaks.
Abb.1: Obwohl Neuron A und Neuron B (AND-Typen) eine identische Funktion besitzen mögen, reagieren sie (bei geringer Leitgeschwindigkeit) nicht gleichartig auf gleiche Impulsbelegungen
Jeffress Verzögerungsmodell 1948
Jeffress soll nach Konishi [2] während eines Forschungsurlaubs 1948 am Caltech eine damals unveröffentlichte Modellschaltung mit nichtideal schnellen Leitbahnen angegeben haben, die aus heutiger Sicht einige wesentliche Elemente biologierelevanter Signalverarbeitung erkennen läßt. Sie bietet einen (bislang allerdings nicht erkannten) Ansatzpunkt zur Interpretation stochastisch verschalteter Netze. Ausgehend von diesem Verzögerungsmodell sollen grundlegende theoretische Bezüge biologischer Informatik dargestellt werden.
Nehmen wir an, die im Bild dargestellte Schallquelle strahlt periodisch kurze Impulse ab. (Im Modell Jeffress werden sinusförmige Zeitfunktionen betrachtet, diese verkomplizieren allerdings das Wesentliche des zu erörternden Vorganges). Dann breiten sich beide Partialwellen in gleichen Zeiten um gleiche Distanzen aus, gleiche Geschwindigkeiten vorausgesetzt. Folglich begegnen sich die Partialimpulse im neuronalen Raum an verschiedenen Stellen, wenn die Quelle ihre Lokalität wechselt. Über den Ort des Eintreffens der Partialimpulse entscheidet einzig die Geschwindigkeit ihres Vorankommens, die Relativität der Ausbreitung der Impulse.
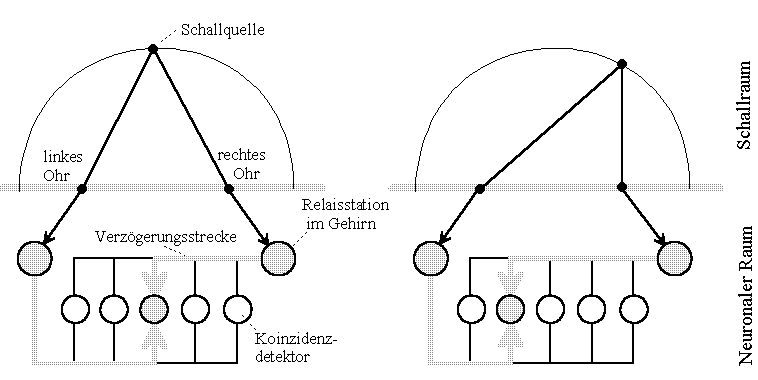
Abb.2: Schallortung der Schleiereule. Modell Lloyd A. Jeffress nach Konishi [2]. Das Schaltungsmodell versucht zu erklären, wie das Gehirn Laufzeitdifferenzen zur Ortung einer Schallquelle benutzen kann. (Zeichnung frei nach im Text bezeichneter Quelle)
Das Modell verdeutlicht, daß unter Berücksichigung endlicher Ausbreitungsgeschwindigkeit verschieden positionierte Schallquellen unterschiedliche Koinzidenzdetektoren (Neuronen) zu maximaler Erregung bringen. Alle anderen Detektoren liefern, je weiter sie vom Interferenzort entfernt sind, schwächer werdende Ausgaben.
Um nun nicht 'Koinzidenzdetektoren' oder andere Konstrukte bemühen zu müssen, die den Vergleich auf korrelierende Zeitfunktionen vorzunehmen gestatten, soll in Analogie zu physikalisch-optischen Auffassungen der Begriff der Interferenz von Zeitfunktionen (Wellen) eingeführt werden. Zwei Impulse oder Wellenzüge mögen an dem Ort in Interferenz stehen, an dem die verschiedenen Wellenanteile gleichzeitig ankommen.
Was bis heute unerkannt blieb, sind die vielfältigen, bereits diesem einfachen Modell zuordenbaren, von gewohnt digitalem abweichenden Eigenschaften:
Erstmalig wird ein Schaltungsmodell angegeben, dessen logische Elemente (Neuronen, Koinzidenzdetektoren) nichtdeterministische Aufgaben erfüllen. Dazu stelle man sich einen Quellort vor, der auf die Mitte zwischen zwei Koinzidenzdetektoren interferiert. Mal wird mehr der rechts liegende, mal mehr der linksliegende Detektor gestreift. Entsprechend funktionieren beide vielleicht nur ab und an. (Diese Aussage ist insofern hochbrisant, als daß sie den prinzipiellen Unterschied zwischen einem interferenziellem Wirkprinzip und den anerkannten Wirkprinzipien digitaler Logikschaltungen ausmacht).
Wird die Schallquelle als Vorlage im Schallraum um ein Stück dx nach einer Seite verschoben, wandert ihr Bild im Neuronenraum proportional um ein anderes Stück -dx' spiegelbildlich in die entgegengesetzte Richtung. Offensichtlich stehen Vorlage und resultierendes Bild in der Proportion dx/dx' zueinander. Für lineare Systeme wird diese Proportion als Abbildungsmaßstab M=dx/dx' eingeführt. Sofern vorausgesetzt werden kann, daß die Vorlage nicht nur aus einem Punkt bestehen kann (und das sei unter gewissen Einschränkungen gegeben), wird die Vorlage homomorph auf das Bild projiziert. Folgend wird - der Optik und Mathematik entlehnt - für solcherart Zusammenhang der Begriff der (neuronalen) Abbildung benutzt.
Die Schärfe der örtlichen Lokalisation ds eines Impulses steht offensichtlich im Zusammenhang mit der geometrischen Länge l des Impulses ds=kl, die geometrische Länge l wiederum ist über die Ausbreitungsgeschwindigkeit mit der zeitlichen Impulsbreite dt verknüpft v=ds/dt=kl/dt.
Die Ausgabe eines Neurons (Koinzidenzdetektors) wird abhängig von den Leitbahnlängen der Eingangsanschlüsse. Dieses Verhalten ist insofern spektakulär, als in digitalen Systemen versucht wird, genau dieses strikt zu vermeiden, um leitungsunabhängige oder verzögerungsunabhängige Funktionalität der Baugruppen zu erreichen.
Die logische Konsequenz aus der funktionalen Längeabhängigkeit der Neuronen ist die, daß damit logische Funktion unabwendbar dem Ort zugeordnet wird, nicht aber dem verknüpfenden Element. Mit anderen Worten: bereits an diesem Modell wird erkennbar, daß u.U. ein Nucleus magnocellularis, operativ um wenige Zentimeter verrückt, nicht mehr dieselbe - also nicht mehr seine eigentliche Aufgabe lösen kann.
Wenn aber logische Funktionalität ortsabhängig wird, dann codiert die Hülle eines Nervengebildes wesentliche, funktionelle Eigenschaften desselben. Folglich beeinflußt bereits die makroskopisch sichtbare, äußere Form eines Nervengebildes dessen Funktionalität.
Damit kommt die logische Funktion nicht mehr dem logischen Element zu, sondern dem Ort, dem sie zugeordnet ist. Unter Maßgabe maximal vernetzter Elemente wird damit der logische Signalfluß nicht mehr durch die Leitbahnen gesteuert, sondern durch die Laufzeit- und Lageproportionen des Raumes. Folglich kann (in gewissen Grenzen) eine stochastische, nichtdeterministische Verdrahtung gewählt werden.
Sollte sich zeigen lassen, daß sich Laufzeitproportionen lageabhängig ändern, so ändern sich damit logische Eigenschaften neuronaler Elemente in Abhängigkeit von der momentanen Stellung oder Lage z.B. eines Körperteils.
Aus dem Modell ist offensichtlich verblüffend viel mehr ablesbar, als bisher bekannt. Unbemerkt hat Jeffress ein erstes Interferenzmodell angegeben, anhand dessen sich bereits wesentliche Eigenschaften neuronaler Netzwerke ableiten lassen, die im krassen Widerspruch zu Eigenschaften bekannter, künstlich-neuronaler Systeme stehen.
(Spätere Anmerkung des Autors: Nach Erscheinen der Interferenz-Netzwerke (IN), wurden die sog. "Neuronal" Networks (NN) umbenannt in "Artifical Neuronal" Networks (ANN).)
Neuronale Abbildung in Eigeninterferenz
Ohne von Jeffress gewusst zu haben, entwickelte der Autor seit 1992 Schaltungen, die die Übertragung von Abbildungen in stochastisch verschalteten Netzen nachzubilden gestatten.

Abb.3: Abbildend wirkende Interferenzschaltung. Ein Generator speist am Ort x1 Impulse gleichzeitig in beide Leitbahnen L ein. Ein multiplizierender Empfänger M liefert nur am Ort der Interferenz x2 eine Ausgabe. Ein Zeiger Z1 wird dabei auf einen Zeiger Z2 abgebildet
Wird der Sender G entlang der x-Richtung bewegt, so muß ein multiplizierender Empfänger M in entgegengesetzter Richtung bewegt werden, um die Möglichkeit des Empfanges zu bekommen. Es handelt sich folglich um eine Schaltung, die abbildend wirkt.
Erweitern wir dieses Schaltungsprinzip, und setzen auf der Sende- und auf der Empfangsseite dicht gepackt unbewegliche Neuronen, so erfolgt dieselbe Abbildung, wenn gesichert wird, daß die verschiedenen Generatoren unkorrelliert zueinander pulsen. Auch darf die Pulsdichte auf den Übertragungsleitungen ein bestimmtes Maß nicht überschreiten, um zu gewährleisten, daß auf der Empfangsseite nichtadressierte Neuronen ansprechen. Wir sehen dann eine funktionell wie strukturell vorzüglich dem Nervenprinzip vergleichbare Schaltung vor uns.
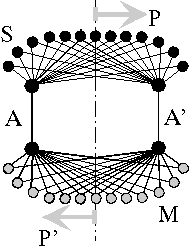
Abb.4: Eindimensionale, neuronale Interferenzschaltung (nach Heinz). Zur Übertragung einer n-dimensionalen Abbildung sind n+1 Kanäle erforderlich: Impulssender S, Multiplizierer M, Übertragungskanäle A, A'; Vorlage P, Bild P'
Die Schaltung verbindet einen Ort P der Geberseite funktionell und spiegelbildlich mit einem Ort P' der Empfängerseite. Interferenz tritt an dem Ort auf, an dem sich die auf verschiedenen Wegen laufenden Impulse treffen, an dem auf allen Wegen 1,2,...m gleiche Laufzeit zwischen Sender (ta) und Empfänger (tb) entsteht.
τ1 = τ2 = ... = τm = tb - ta
Die Größe zusammengehöriger Orte dx ist zur geometrischen Impulsbreite λ = vτ mit einem Faktor k proportional, dx = kλ. Wird der Ort der Interferenz durch Laufzeitveränderungen der Leitbahnen verschoben, übernehmen andere Logikelemente die Aufgabe bisheriger. Die Abbildung arbeitet solange störungsfrei, solange der Abstand a = v(tj - ti) zweier Folgeimpulse größer als die Länge L des Feldes ist, v(tj - ti) > L.

Abb.5: Interferenz auf einem Feld mit vier speisenden Kanälen (Momentaufnahme). a) Zeitliche Relation der ankommenden Impulse, b) Bestimmung eines möglichen Interferenzortes
Fremdinterferenz-Abbildungen (Allgemeine Interferenz-Abb.)
Neben der Interferenz eines Impulses mit sich selbst in Eigeninterferenz ist es möglich, daß ein Impuls in Interferenz zu irgendeinem Vorgänger- oder Folgeimpuls tritt. Dieser Typus möge als Fremdinterferenz bezeichnet werden.

Abb.6: Neuronale Beugung als Interferenz verschiedener Impulse miteinander. Der Abstand zwischen den Maxima korrespondiert mit dem Pulsabstand (j-k)
Benachbarte Maxima besitzen gerade eine Wegdifferenz s in der Leitbahnlänge, die dem zeitlichen Abstand Τ = (tj - ti) zweier oder vielfacher Folgeimpulse entspricht,
s = v (tj - ti) = vΤ.
Proportionalitätskonstante ist die Leitgeschwindigkeit v. Entsprechend gilt für Interferenz zweier Leitbahnen zwischen Quelle und Interferenzort zusätzlich zur Eigeninterferenz
τi + nΤ = τj + mΤ mit (n,m) ∈ N
woraus für k = n - m folgt τj - τi = kΤ. Für mehrfache Übertragungen entstehen die Interferenzorte
τ1 - a1Τ = τ2 - a2Τ = ... = τn - anΤ mit aν ∈ N.
darin ist die Lösung für Eigeninterferenz mit a1 ... an = 0 bzw. für Τ > τν enthalten. Folglich entstehen fehlerfreie Abbildungen nur bei einer schwachen Kanalbelegung! Überlastete Kanalbelegungen, zB. für Τ < τν können Abbildungsfehler (Schmerz?) erzeugen.
(Spätere Anmerkung d.A.: Innervationsgebiete von Hautnerven besitzen eine Ausdehnung von mehreren Hundert Quadratmillimetern. Demgegenüber lassen sich Erregungen im Millimeterabstand wahrnehmen. Wie ist das möglich?)
Mit dem Elektro-Enzephalographen (EEG) sollen eine Mehrfachausbreitung und eine relative Verschiebung einer Wellenfront in Bezug auf den Empfänger nachgewiesen werden. Dazu werden in den Daumen (anatomisches Gebiet des N. radialis und des N. medianus) mit einem Paar Ringelektroden Stromimpulse (Peak 4mA, Dauer 1ms) eingespeist, die oberhalb des Handgelenks an Stellen, an denen diese Nerven nahe an die Oberfläche kommen, mittels Oberflächenelektroden abgegriffen werden können (siehe Foto). Die an zwei Paar Elektroden abgegriffenen Spannungsverläufe von N. medianus und N. radialis sind im Bild dargestellt, oben N. radialis, unten N. medianus.
Bei einer Veränderung der Stellung des Daumens zwischen den Endlagen verändert sich die Lage der Impulse gegeneinander. Zur Lagevariation sind zwei Messungen erforderlich. In einer ersten Messung wird der Daumen auswärts gebeugt. Es wird eine relative Verzögerungszeit τ1 zwischen den Abgriffen für N. meridianus und N. radialis ermittelt. Im zweiten Experiment wird der Daumen einwärts gebeugt. Es entsteht eine kleinere, je nach Lage der Abgriffe zueinander sogar im Vorzeichen entgegengesetzte, relative Verzögerungszeit τ2. Während des Versuchs darf die Lage der Elektroden unter keinen Umständen verändert werden, sämtliche Elektroden sind sorgfältigst zu fixieren, um ein Verrutschen zu vermeiden.
 b)
b)

Abb.7: Meßanordnung. Die erregenden Ringelektroden am Daumen und die Ableitelektroden an N. medianus und N. radialis sind zu erkennen. Das Armband stellt eine Erdungselektrode dar, um Fremdeinflusse (Artefakte) zu vermindern.

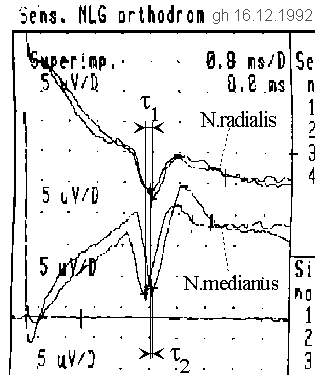
Abb.8: Daumen-Experiment. Wird der Daumen in verschiedener Richtung fixiert, sind relativ zueinander verschiedene Verzögerungszeiten messbar
Im EMG-Auszug werden beide Messungen übereinander gelegt, um die Differenz im Maximum entsprechend deutlich vergleichen zu können. Versehentlich wurde ein Ergebnis ausgedruckt, bei dem unbemerkt im Experiment ein geringfügig differierender Erregerstrom eingestellt war. Daraus erklären sich die Abweichungen im Verlauf der Medianus- Kurven. Aufgrund der relativ unkomplizierten Nachvollziehbarkeit dieses Experiments fiel die Entscheidung leicht, dennoch die historisch originalen, in der Erregung der Stunde vertauschten Originale des Erstversuches am 16.12.1992 zu veröffentlichen, da auch sie das wesentliche Ergebnis wiedergeben.
Die Impulse werden mit einem Durchschnittsbildner (Averager) aus dem Rauschen zufälliger, anderer Impulse geholt. Nach etwa 10 Zyklen ist die abgebildete Darstellungsqualität erreicht. Die Verschiebung ist am N. medianus besonders deutlich zu erkennen. Der jeweils größere Biegeradius benötigt eine längere Laufzeit. Aufgrund der Wahl der Polarität der Elektroden liegt die Impulsspitze nach unten. Die Impulse sind in der Mitte etwa 1ms breit (Darstellung: 0,8 ms/Raster vertikal, 5 μV/Raster horizontal), wobei eine nicht näher quantifizierbare Mittelung durch den Averager eine zusätzliche Verschleifung der Impulsform verursacht. Deshalb ist anzunehmen, daß die tatsächliche Impulsform wesentlich steiler ist.
Es ist zu erkennen, daß sich die Laufzeitdifferenz zwischen den beiden diametralen Beugestellungen des Daumens verändert. Auswärts gebeugt entsteht eine Verzögerung von
τ1 = τr - τm = -0.3ms,
einwärts gebeugt ist eine Verzögerung von
τ2 = τ'r - τ'm = +0.2ms
zwischen N. radialis und N. medianus zu messen. In der Handskizze sind symbolisch Wellenfronten eingetragen, die diesen Sachverhalt verdeutlichen. Die Entfernung zwischen Erregung und Abgriff betrug etwa 13 cm, die durchschnittliche Leitgeschwindigkeit v = 32,5 m/s. Die Absolutzeitdifferenz zwischen den Orten zweier Neuronen x und y, die potentiell die voneinander verschiedenen Laufzeitdifferenzen mit maximaler Erregung empfangen und auswerten könnten, beträgt
τ = τ2 - τ1 = 0.5ms.
Folglich läßt sich eine Differenz in der Länge der Zuführungen zu irgendwo mit maximaler Wahrscheinlichkeit empfangenden Neuronen bei willkürlich konstant angenommener Leitgeschwindigkeit v = 32,5 m/s bestimmen zu
s = v τ = 32.5 m/s * 0.5 ms = 16.25 mm.
Ist die natürliche Impulsbreite kleiner als 0,5ms (sie ist vermutlich wesentlich kleiner als die vom Averager aufgezeichnete), so findet eine aus dem Daumen stammende Erregung unterschiedliche Zieladressen in Abhängigkeit vom Grad der Beugung.
Vernetzungsalgorithmus für Interferenzabbildungen
Eine Abbildung zwischen Sender S und Empfänger M entsteht an der Stelle, an der die Laufzeiten eines Impulspaars über die Übertragungsleitungen A und A' identisch sind. Es wird deutlich, daß der Algorithmus zur Gestaltung von Verbindungen einfacher ist, als beispielsweise die Verlegung einer Hausklingelanlage entsprechender Kanalkapazität.
Vernetzungsalgorithmus für Interferenzabbildungen (mono- und bidirektional), siehe Abb.4:
Randbedingungen sind die, daß Orte, auf die Sender bzw. Empfänger lokalisiert sind, strengen Zeitbedingungen unterliegen, und daß die Sender asynchron zueinander pulsen. Interferenzen zwischen Orten müssen möglich sein. Damit wird eine Hüllen- und Körpergeometrie definiert.
Es wird deutlich, daß zum Zustandekommen einer fehlerfreien Puls- Interferenzabbildung nicht einmal ein Verdrahtungsplan (zB. wie ihn ein Elektriker zur Verlegung einer Hausklingelanlage benutzt), vonnöten ist.
Untersuchungen zur Relativität neuronaler Impulsausbreitung und zu Interferenzerscheinungen geometrisch kurzer Impulse in neuronalen Netzen lassen eine kausale Bindung neuronaler Kommunikation, Adressierung und Speicherung an neuronale Laufzeiträume erkennen. Biologische, neuronale Systeme sind durch schmale Impulsbreiten und langsame Leitgeschwindigkeiten charakterisiert.
Die geometrische Impulslänge ist klein im Verhältnis zu den Abmessungen des Systems (Körpers). Die Impulslänge ist meßbar. Der Nachweis dafür ist mit einem herkömmlichen EEG- Gerät möglich. Biologische Systeme sind lernfähig, aufgrund des vor dem Lernen unbekannten Zielortes müssen Impulse diffus in verschiedene Leitbahnen gestreut werden. Sind verschiedene Nervenzellen in unterschiedlicher Laufzeit-Geometrie an die impulstragendenen Nervenstränge angeschlossen, wird die Nervenzelle maximal erregt, die in bestmöglicher Interferenz zum Sendeort liegt, dh. bei der alle Partialimpulse gleichzeitig eintreffen.
Folglich ist damit eine Adressierung neuronaler Information unter Berücksichtigung diffuser Impulsausstreuung und diffuser Verschaltung gegeben. Ein hypotetisches Rechenbeispiel illustriert diesen Sachverhalt. Konstruktionsregeln für interferenzielle Abbildungen werden erörtert. Sie sind einfacher und damit im Sinne genetischer Strukturevolution codeleichter, als die vergleichbarer Klingeldrahtmodelle. Laufzeiteffekte, die als Relativität von Laufzeiten in Neuronen und auf Leitbahnen erscheinen, stellen das Adressierungsverfahren neuronaler Information an sich dar. So kann über sie das Zielgebiet einer zu übertragenden Information in Größe und Lage gesteuert werden.
Die Untersuchung kann als Nachweis gewertet werden, daß biologische, neuronale Systeme durch Zustandsmaschinen-Modellierung der Art nach McCulloch/Pitts unter Vernachlässigung der Laufzeit und der geometrischen Länge neuronaler Impulse im Ansatz nicht zureichend modelliert werden können. Insofern wird es verständlich, daß die großen Erfolge der künstlichen Intelligenzforschung bisher wenig zum Verständnis biologischer Systeme beitragen konnten. Ein mit den Untersuchungen entstandenes Manuskript [3] wird in Buchform erscheinen.
Mein herzlicher Dank gilt dem EEG-Spezialisten Herrn Dr. Torsten Griepentrog für die freundliche Unterstützung bei der Aufnahme des EMG in der Landesklinik Teupitz (Land Brandenburg). Herzlichen Dank für das freundliche Entgegenkommen auch an den Chefarzt für Psychiatrie, Dr. Alfons Horn.
[1] Schmidt, R.F., Thews, G.: Physiologie des Menschen, 24. Aufl., Springer-Verlag Berlin, 1990, S. 40
[2] Konishi, M.: Die Schallortung der Schleiereule. Spektrum der Wissenschaft, Juni 1993 S.58-71
[3] Heinz, G.: Neuronale Interferenzen. GFaI-Report vom 23.12.1993 (300 S.)
[4] Heinz, G.: Modelling Inherent Communication Principles of Biological Pulse Networks. SAMS 1994, Vol.15, No.1, Gordon & Breach Science Publ., Printed in the USA.
[5] Heinz, G.: Neuronale Interferenzen. Öffentliche Vorträge:
- GFaI/GMD-FIRST im Hause 13.7, Akademiegelände Berlin- Adlershof am 27.8.93
- Fachhochschule für Technik und Wirtschaft Berlin-Karlshorst FB3 (Prof. Matschke), 28.5.93
- Öffentlicher Vortrag an der Technischen Fachhochschule Berlin-Wedding, Labor für Künstliche Intelligenz (Prof. Hamann) am 27.10.93
- Öffentlicher Vortrag im Institut für Mikroelektronik der Technischen Universität Berlin am 27.1.94
Author
Dr. Heinz, GFaI e.V., Rudower Chaussee 5, Haus 13.7, 12484 Berlin,
Tel. +49-30-6392-1624, Fax -1602, e-mail heinz@gfai.fta-berlin.de
aktualisiert:
e-mail info@gheinz.de
URL http://www.gheinz.de
Visitors since Dec. 2021: